
Roger Schneiter
PER 07 - 0.307
+41 26 300 8654
E-mail
Our lab is working on lipid metabolism, using Baker’s yeast as a unicellular model organism. We are looking at two different questions to understand how cells regulate and adapt their lipid composition to changes in environmental conditions to maintain a homeostatic balance:
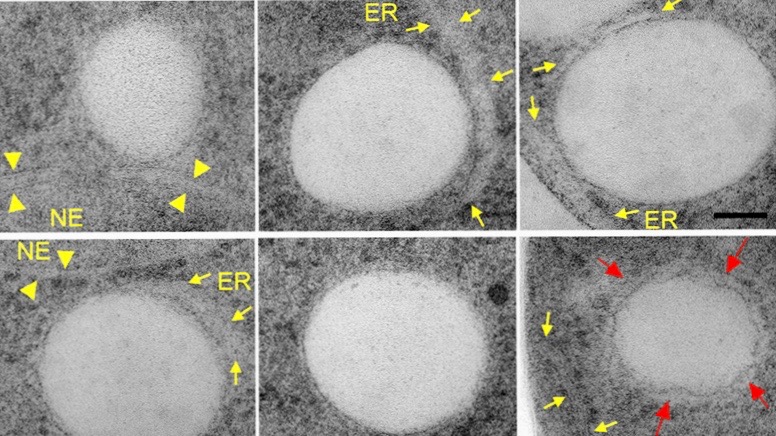
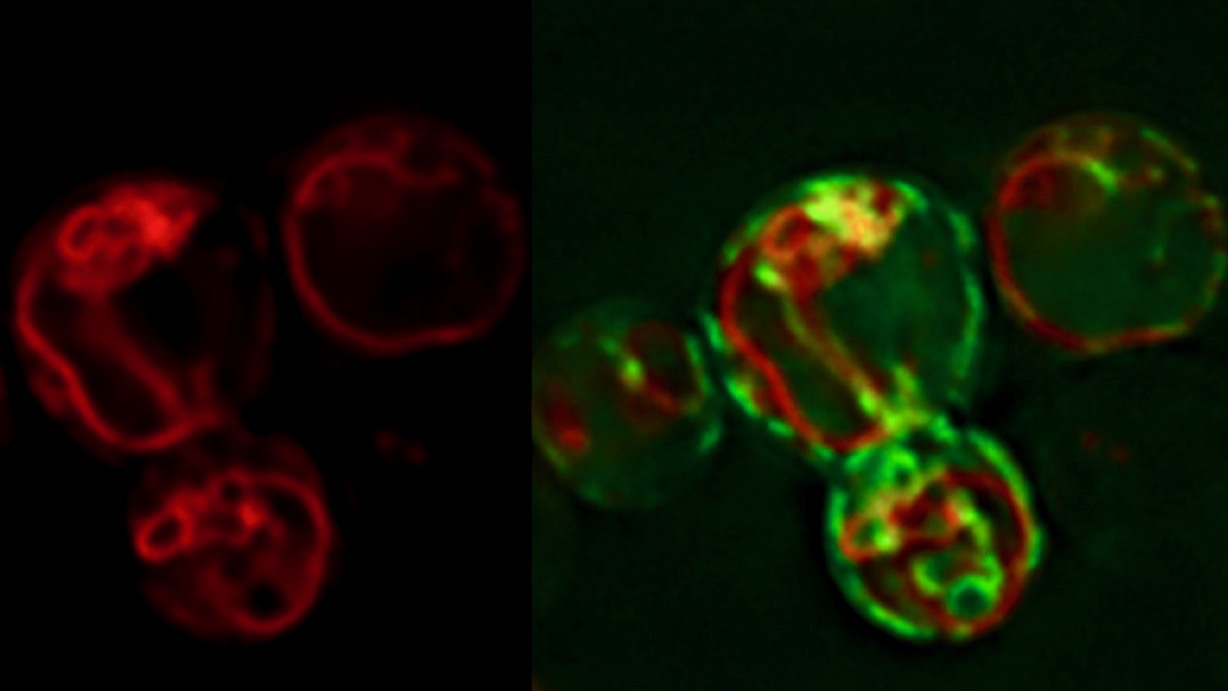
Lipid droplets are intracellular globular shaped structures that serve to store metabolic energy in form of neutral lipids (fat). They dynamically form and grow in size under conditions of energy excess and they shrink upon energy demand. Lipid droplets are composed of a hydrophobic core made of triacylglycerol and steryl esters. This neutral lipid core is covered by a phospholipid monolayer and harbors a set of proteins that are specifically targeted to the lipid droplet surface. While yeast cells typically harbor up to a dozen of these droplets, adipocytes are filled with one large lipid droplet. As the core of the lipid droplet is highly hydrophobic and not soluble in the aqueous environment, the synthesis, transport and storage of these lipids has to be orchestrated with the biogenesis of lipid droplets. We are using live cell microscopy to understand how these processes are coordinated and where exactly they occur in the cell. Our data indicate that lipid droplets are in close proximity to the endoplasmic reticulum (ER) allowing for rapid and bidirectional exchange of both lipids and proteins between the ER membrane and the lipid droplets.
In our recent study, we show that the biogenesis of LDs occurs at discrete ER subdomains defined by two membrane proteins, Fld1 and Nem1. Fld1 (seipin in human) localizes to ER-LD contact sites and fulfills a key function in LD biogenesis. Nem1, on the other hand, is part of a protein phosphatase complex that activates Pah1, a key enzyme for the production of DAG. Remarkably, Fld1 and Nem1 colocalize together at ER subdomains to recruit the TAG-synthases, Lro1 or Dga1, as well as additional factors that promote LD biogenesis. This highly localized formation of TAG is important, because in cells lacking either Fld1 or Nem1, TAG synthesis occurs ectopically throughout the ER. Such ectopically formed LDs do not contain the complete set of LD proteins and hence are impaired in function. Based on these findings, we propose a model for a stepwise initiation of LD biogenesis and a spatially and temporally ordered recruitment of proteins and lipids to ensure a regulated biogenesis of functional LDs.
See the SPOTLIGHT commentary article in JCB https://doi.org/10.1083/jcb.202006025
 Model of LD biogenesis from specialized ER subdomains. Subdomains of the endoplasmic reticulum (ER) are first established by the colocalization of seipin (Fld1) and Nem1, an activator of diacylglycerol (DAG) production. These Fld1/Nem1-sites then recruit additional factors such as Yft2 and Pex30 and they become enriched in DAG. Sites containing Fld1/Nem1/Yft2/Pex30 and DAG then recruit the neutral lipid synthesizing enzymes Lro1 and Dga1. These enzymes convert DAG into triacylglycerol (TAG) thereby promoting a highly localized biogenesis of lipid droplets (LDs). Newly formed LDs become stabilized by Pet10, a perilipin homolog and LD scaffolding factor, and recruit Erg6 and other bona fide LD proteins.
Model of LD biogenesis from specialized ER subdomains. Subdomains of the endoplasmic reticulum (ER) are first established by the colocalization of seipin (Fld1) and Nem1, an activator of diacylglycerol (DAG) production. These Fld1/Nem1-sites then recruit additional factors such as Yft2 and Pex30 and they become enriched in DAG. Sites containing Fld1/Nem1/Yft2/Pex30 and DAG then recruit the neutral lipid synthesizing enzymes Lro1 and Dga1. These enzymes convert DAG into triacylglycerol (TAG) thereby promoting a highly localized biogenesis of lipid droplets (LDs). Newly formed LDs become stabilized by Pet10, a perilipin homolog and LD scaffolding factor, and recruit Erg6 and other bona fide LD proteins.
Key Publications:
Choudhary V, El Atab O, Mizzon G, Prinz WA & Schneiter R (2020). Seipin and Nem1 establish discrete ER subdomains to initiate yeast lipid droplet biogenesis. J. Cell Biol. 219, DOI: 1083/jcb.201910177
Mishra S, Khaddaj R, Cottier S, Stradalova V, Jacob C & Schneiter R (2016). Mature lipid droplets are accessible to ER luminal protein. J. Cell Sci. 129: 3803-3815.
Jacquier N, Choudhary V, Mari M, Toulmay A, Reggiori F & Schneiter R (2011). Lipid droplets are functionally connected to the endoplasmic reticulum in Saccharomyces cerevisiae. J. Cell. Sci. 124: 2424-2437.
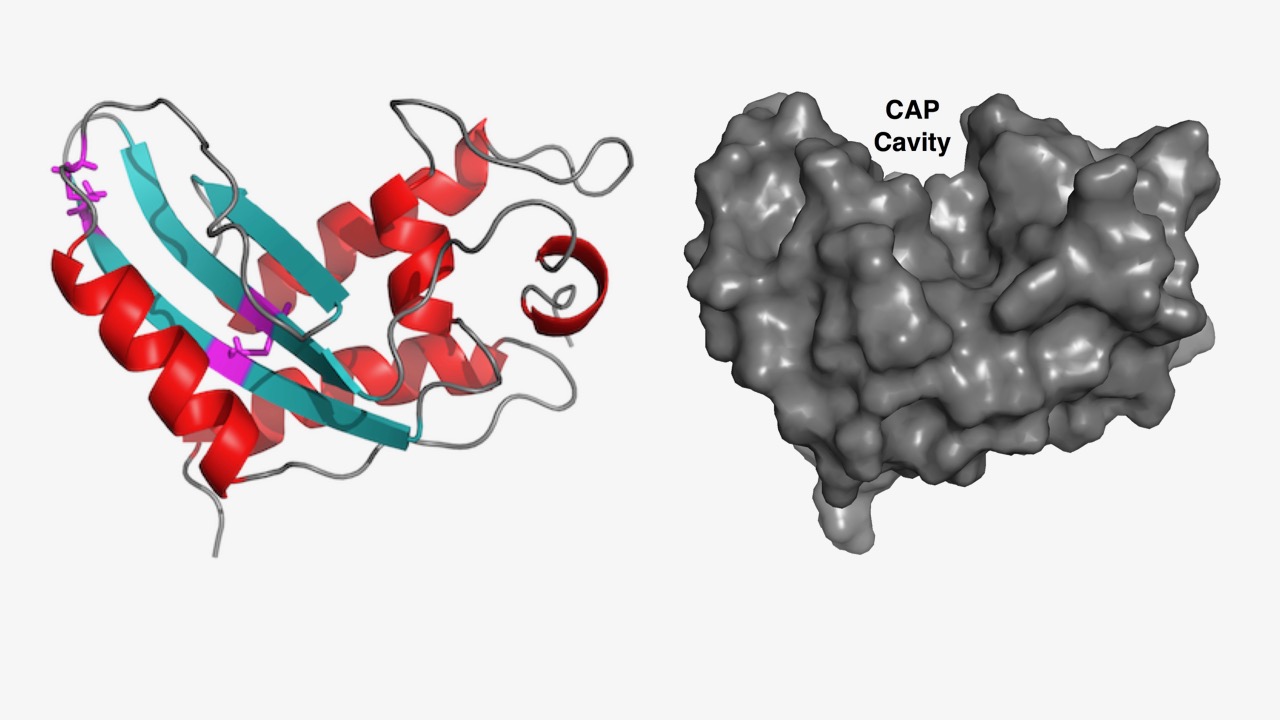
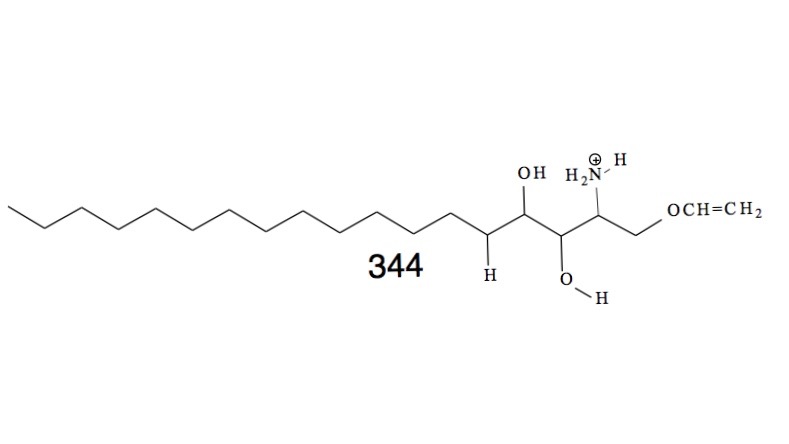
Proteins belonging to the CAP superfamily are found in all kingdoms of life (Pfam PF00188). This superfamily was named after three founding members: Cysteine-rich secretory proteins (CRISP) found in mammals, antigen 5 (Ag5) in stinging insects and pathogenesis-related protein 1 (PR-1) in plants. The human genome encodes for 32 family members, whereas yeast has 3 CAP members, termed Pathogen Related in Yeast, Pry1-3. These proteins have been implicated in many different physiological processes ranging from immune defense in mammals and plants, sperm maturation and fertilization, prostate and brain cancer, pathogen virulence, and venom toxicity. They are most often secreted glycoproteins that adapt a unique alpha-beta-alpha sandwich fold. We found that these proteins bind both sterols and fatty acids in two independent lipid binding sites and they promote the export of these lipids in vivo. We are now testing whether the ability of these proteins to sequester lipids can account for their physiological functions reported for different organism.
 Structural model of a possible open-loop conformation of the yeast Pry1 protein that could accommodate small hydrophobic molecules such as cholesterol. For illustration purposes, the cholesterol molecule (green) was manually positioned so as to fill optimally the open cavity.
Structural model of a possible open-loop conformation of the yeast Pry1 protein that could accommodate small hydrophobic molecules such as cholesterol. For illustration purposes, the cholesterol molecule (green) was manually positioned so as to fill optimally the open cavity.
Key Publications:
Cottier S, Darwiche R, Meyenhofer F, Debelyy MO & Schneiter R (2020). The yeast cell wall protein Pry3 inhibits mating through highly conserved residues within the CAP domain. Biol. Open. DOI: 1242/bio.053470
Darwiche R, Mène-Saffrané L, Gfeller D, Asojo OA & Schneiter R. (2017). The pathogen-related yeast protein Pry1, a member of the CAP protein superfamily, is a fatty acid-binding protein. J. Biol. Chem. 292: 8304-8314.
Choudhary V & Schneiter R (2012). Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. PNAS 109: 16882-16887.







Alumni:
PER 07 - 0.307
+41 26 300 8654
E-mail